血脑屏障是位于血液和脑脊液、血液和脑细胞之间,由脑毛细血管壁、内皮细胞和神经胶质细胞等形成的天然屏障。由于血脑屏障主要由内皮细胞间紧密连接形成,大多数大分子物质(包括蛋白类药物)无法穿越,因此这类物质进入中枢神经系统(central nervous system,CNS)的效率受到严重限制。传统的针对CNS的给药方式,包括颅内给药、鞘内给药、脑室内给药和化学破坏血脑屏障给药等,具有侵入性,可能对脑组织造成损伤,不利于重复给药。脑组织暴露不足和给药方式的限制,使许多针对CNS的抗体药物无法发挥应有的药效。 近日,来自Denali Therapeutics Inc.的科学家在《Science Translational Medicine》上介绍了他们设计的双特异性抗体,称之为血脑屏障运输工具抗体(antibody transport vehicle,ATV)。这种抗体被认为可特异性地与大脑内皮细胞表面上表达的RMT受体结合,利用受体介导的胞吞效应(receptor-mediated transcytosis,RMT)穿过血脑屏障,极大地提高抗体药物向中枢神经系统的传递效率。
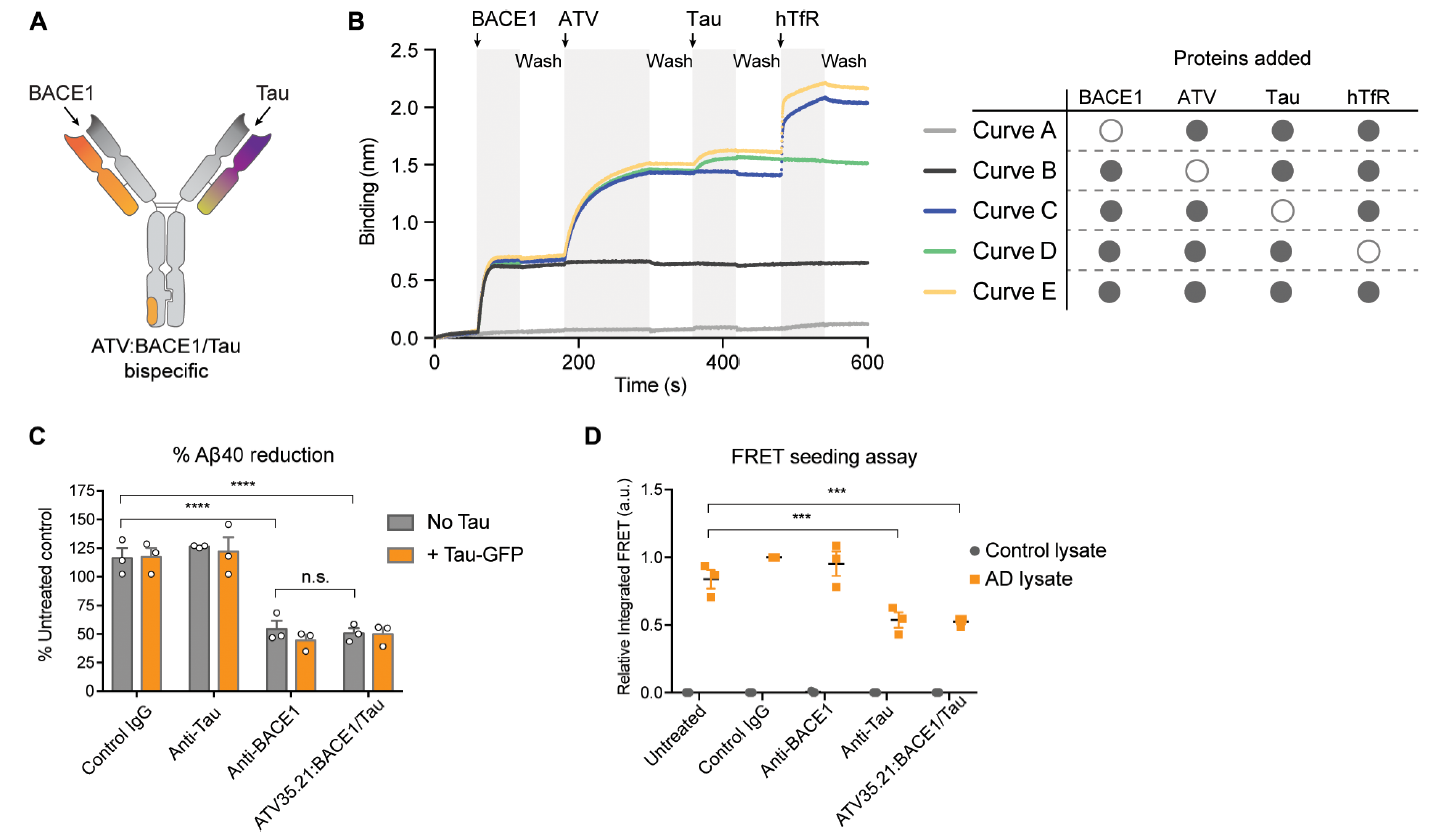
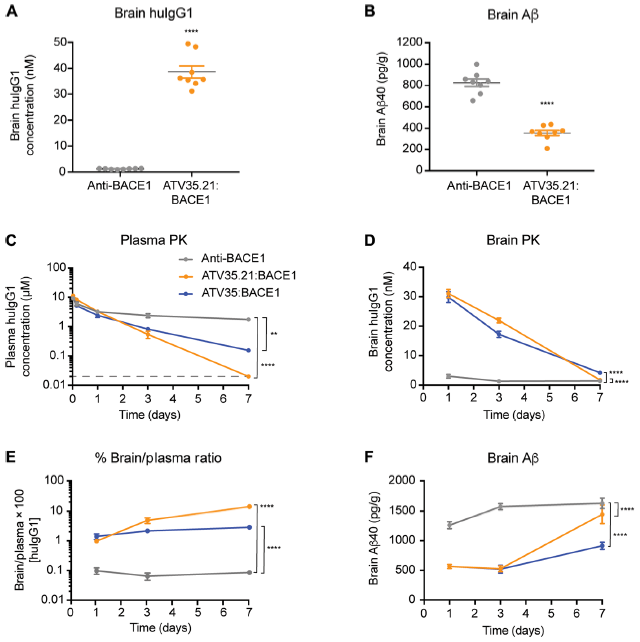
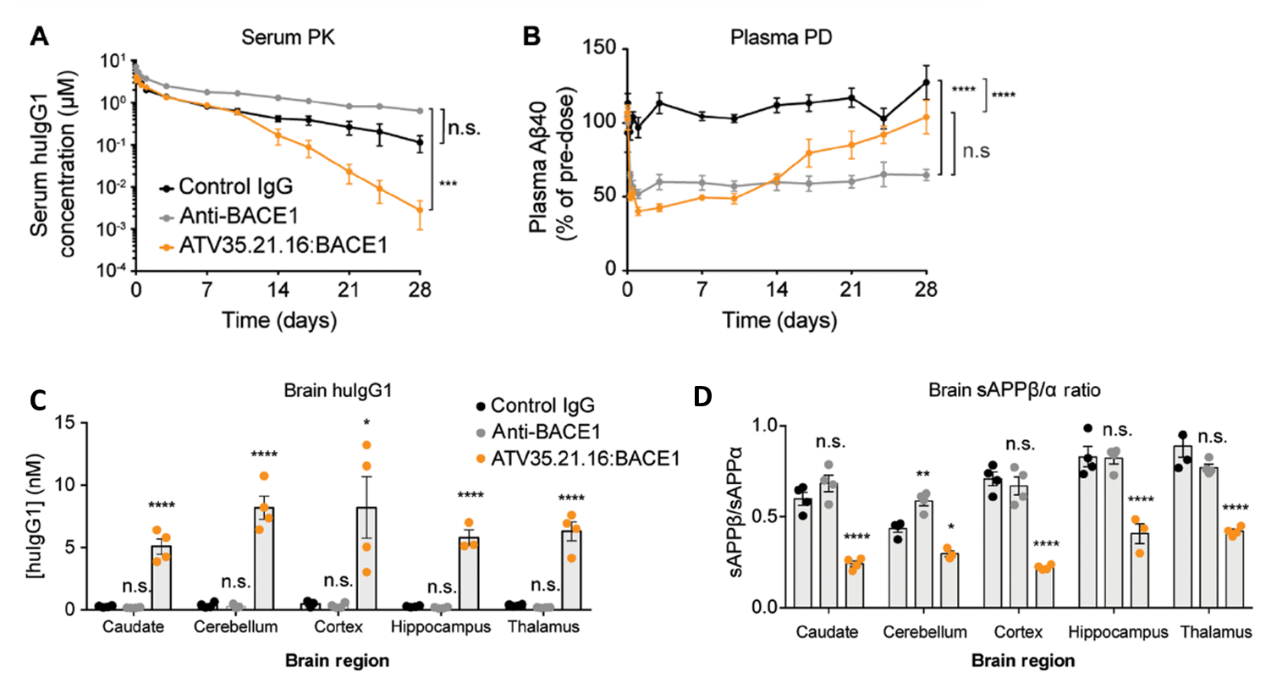
ATV靶点的选择 这项研究中选择了转铁蛋白受体(transferrin receptor,TfR)作为穿越血脑屏障的靶点。TfR在脑内皮细胞上高度表达,是重要的配体非依赖性胞吞的受体,早期动物模型实验结果显示,与TfR单价结合的低亲和力的抗体可以提高大脑对药物的摄取能力。大脑和脑脊液中的β淀粉样蛋白(β-secretase 1,BACE1)可作为阿尔兹海默症的生物标志物,前期数据表明抗BACE1抗体可减少小鼠大脑中β淀粉样蛋白的含量,因此研究人员选择了BACE1作为ATV的治疗靶点。 ATV的构建和亲和力改造 研究人员在人IgG1的Fc端选取了一段既远离FcRn和FcɣR结合位点又适合作为新的抗原结合位点的9个氨基酸组成的序列(图1A)。对这个序列进行随机化重组,然后利用酵母展示技术和流式细胞分选技术,筛选出了四个与hTfR胞外段结合的Fc蛋白(图1B)。将这四个Fc蛋白与抗BACE1的Fab融合构建出ATV:BACE1重组抗体。这些ATV:BACE1抗体在蛋白水平(图1C,ELISA)和细胞水平(图1D,免疫荧光染色)都能与hTfR结合。最后在这四个重组抗体的基础上,通过文库筛选获得了亲和力较高且能与食蟹猴TfR(cTfR)结合的一系列ATV:BACE1抗体(图2)用于后续体内外功能验证。 图1,A野生型Fc同源二聚体蛋白结构模型,橙色部位为筛选的9个氨基酸所处位置;B,筛选得出的4个Fc序列(TV);C,ATV:BACE1重组抗体与hTfR蛋白的结合能力;D,ATVc:BACE1与HEK293细胞(表达TfR,不表达BACE1)的结合能力。 图2:不同ATV:BACE1克隆对人TfR、食蟹猴TfR的亲和力。 ATV:BACE1在细胞水平上的内化 研究人员使用knob-in-hole技术合成了Fc上具有TfR单个结合位点的ATV:BACE1异源二聚体(图3A)。将ATV:BACE1异源二聚体与HEK293细胞(表达TfR)或CHO细胞(不表达TfR)在37℃共孵育30 min后,使用抗人IgG荧光染料染色,观察到HEK293细胞内化的荧光标记数与ATV:BACE1浓度呈剂量依赖效应(图3B),而与ATV:BACE1共孵育的CHO细胞及与Anti-BACE1共孵育的HEK293细胞均无荧光标记(图3B)。通过对内化的TfR荧光染色,显示约有50%的ATV:BACE1与TfR定位在细胞内相同的位置。这些结果表明ATV:BACE1在细胞上的内化依赖于TfR,且ATV:BACE1与TfR的结合不影响TfR的内化。 图3:A,knob-in-hole型ATV:BACE1异源二聚体的结构;B,HEK293及CHO细胞中ATV:BACE1的染色;C,HEK293细胞中anti-BACE1的染色;D,HEK293细胞中ATV:BACE1与TfR的共染色。 双特异性ATV在细胞水平上的疗效 为了使ATV发挥更好的神经退行性疾病疗效,研究人员将针对两个直接与阿尔兹海默症形成有关靶点(β-secretase、Tau蛋白)的片段与ATV融合,制备了ATV:BACE1/Tau双特异性抗体(图4A),这种双特异性抗体可以同时与BACE1、Tau、hTfR三个靶点结合(图4B)。与同型对照IgG和anti-Tau抗体相比,ATV:BACE1/Tau双特异性抗体减少了50%人初级皮质神经元细胞中Aβ-secretase 40的数量(图4C)。荧光共振能量转移(fluorescence resonance energy transfer,FRET)的实验结果显示,预先接种Tau蛋白的HEK293细胞经ATV:BACE1/Tau处理后能有效降低细胞中Tau蛋白的聚集(图4D)。这些结果表明ATV:BACE1/Tau在细胞水平上具有良好的生物治疗活性。 图4:A,ATV:BACE1/Tau双特异性抗体的结构;B,ATV:BACE1/Tau与BACE1、Tau、hTfR结合的生物干涉结果;C,ATV:BACE1/Tau处理后人神经细胞中Aβ40的含量;D,显ATV:BACE1/Tau处理后预先接种Tau蛋白的HEK293细胞中Tau蛋白的聚集情况(荧光共振能量转移)。 ATV:BACE1在啮齿类及非人灵长类动物模型上的疗效 TfRmu/hu转基因小鼠和食蟹猴模型中,ATV:BACE1表现出与亲和力正相关的药代动力学数据与生物清除效率(图5C,D,图6A)。TfRmu/hu转基因小鼠单次给药24小时后(50mg/kg),与anti-BACE1治疗组小鼠相比,ATV:BACE1治疗组小鼠增加了40倍的药物脑部暴露率(图5A),同时可减少57%脑部可溶性Aβ40的含量(图5B)。共聚焦显微镜下大脑皮层染色结果进一步表明ATV:BACE1给药组小鼠大脑神经细胞中有显著的药物吸收(图5G,H)。单次给药后,脑部药物暴露率的增强可持续7天以上,持续维持脑部较低水平可溶性Aβ40含量(图5E,F)。食蟹猴模型实验结果显示,在静脉单次给药(30 mg/kg)48h后,ATV:BACE1治疗组食蟹猴在尾状核、小脑、脑皮层、海马区、丘脑等脑区ATV:BACE1治疗组药物浓度较control IgG对照组及anti-BACE1组均有显著增加(图6C),且这些区域内可溶性淀粉样前体蛋白β的含量减少了32%-69%(图6D)。以上实验结果在动物模型上证明了ATV:BACE1具有良好的穿越血脑屏障能力,以及治疗阿尔兹海默症等神经退行性疾病的潜在效果。 图5:A,TfRmu/hu转基因小鼠单次给药24h后脑部hIgG的浓度;B,TfRmu/hu转基因小鼠单次给药24h后脑部Aβ40的数量;C,TfRmu/hu转基因小鼠血清中药物PK数据(0-7天);D,TfRmu/hu转基因小鼠脑部药物PK数据(0-7天);G、H,共聚焦显微镜TfRmu/hu转基因小鼠神经免疫荧光染色结果。 图6:A,ATV:BACE1在食蟹猴血清中PK数据;B,ATV:BACE1治疗后Aβ40在食蟹猴血浆中PD数据;C,ATV:BACE1在食蟹猴尾状核、小脑、脑皮层、海马区、丘脑等脑区的浓度;D,ATV:BACE1治疗后食蟹猴尾状核、小脑、脑皮层、海马区、丘脑等脑区APPβ的含量。 尽管过去30年间抗体药物已在癌症以及自身免疫疾病治疗领域发挥至关重要的作用,在神经退行性疾病治疗上的应用却止步不前。如何增强抗体药物脑部的吸收效率是该领域的痛点。该文通过构建具有RMT受体结合位点的双特异性抗体为增强抗体药物脑部暴露率提供了新思路,相信未来这一理念有望解决神经退行性疾病治疗的难题。 原创内容未经授权,禁止转载至其他平台。
参考文献 Kariolis, M. S., Wells, R. C., Getz, J. A., Kwan, W., Mahon, C. S., Tong, R., ... & Assimon, V. A. (2020). Brain delivery of therapeutic proteins using an Fc fragment blood-brain barrier transport vehicle in mice and monkeys. Science Translational Medicine, 12(545). |